Содержание
Ликвор — это спинномозговая жидкость со сложной физиологией, а также механизмами образования и резорбции.
Она является предметом изучения такой науки, как ликворология.
Единая гомеостатическая система контролирует спинномозговую жидкость, окружающую нервы и глиальные клетки в мозгу, и поддерживает относительное постоянство ее химического состава в сравнении с химическим составом крови.
Внутри мозга находятся три вида жидкости:
- кровь, которая циркулирует в обширной сети капилляров;
- ликвор — спинномозговая жидкость;
- жидкость межклеточных пространств, которые имеют ширину около 20 нм и свободно открыты для диффузии некоторых ионов и крупных молекул. Это главные каналы, через которые питательные вещества достигают нейронов и глиальных клеток.
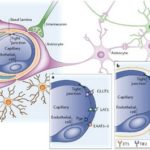
Гомеостатический контроль обеспечивается эндотелиальными клетками мозговых капилляров, эпителиальными клетками сосудистых сплетений и арахноидальными мембранами. Связь ликвора можно представить следующим образом (смотрите схему).
 и структур головного мозга")
Резервуары ликвора связаны:
- с кровью (непосредственно через сплетения, арахноидальную оболочку и т.д., а косвенно через гематоэнцефалический барьер (ГЭБ) и экстрацеллюлярную жидкость мозга);
- с нейронами и глией (косвенно через внеклеточную жидкость, эпендиму и мягкую мозговую оболочку, а непосредственно — в некоторых местах, особенно в III желудочке).
Образование ликвора (спинномозговой жидкости)
Ames и соавторы исследовали извлеченную жидкость из сосудистых сплетений. Результаты, полученные авторами, еще раз доказали, что сосудистые сплетения боковых, III и IV желудочков являются основным местом образования ликвора (от 60 до 80%). Спинномозговая жидкость может возникать также в других местах, о чем предполагал еще Weed. В последнее время это мнение подтверждается новыми данными. Однако количество такого ликвора значительно больше, чем образованного в сосудистых сплетениях. Собрано достаточно доказательств, подтверждающих образование спинномозговой жидкости вне сосудистых сплетений. Около 30%, а по данным некоторых авторов, и до 60% ликвора возникает вне сосудистых сплетений, но точное место его образования остается предметом дискуссий. Ингибирование фермента карбоангидразы ацетазоламидом в 100% случаев прекращает образование ликвора в изолированных сплетениях, но in vivo его эффективность снижается до 50-60%. Последнее обстоятельство, как и исключение ликворообразования в сплетениях, подтверждают возможность появления спинномозговой жидкости вне сосудистых сплетений. Вне сплетений ликвор образуется в основном в трех местах: в пиальных кровеносных сосудах, эпендимальных клетках и мозговой интерстициальной жидкости. Участие эпендимы, вероятно, незначительно, о чем свидетельствует ее морфологическая структура. Главным источником образования ликвора вне сплетений служит мозговая паренхима с ее капиллярным эндотелием, который образует около 10—12 % спинномозговой жидкости. Для подтверждения этого предположения изучались внеклеточные маркеры, которые после их введения в мозг обнаруживались в желудочках и подпаутинном пространстве. Они проникали в эти пространства независимо от массы их молекул. Сам эндотелий богат митохондриями, что свидетельствует об активном метаболизме с образованием энергии, которая необходима для этого процесса. Экстрахориоидальной секрецией объясняется и отсутствие успеха при сосудистой плексусектомии при гидроцефалии. Наблюдается проникновение жидкости из капилляров непосредственно в вентрикулярное, субарахноидальное и межклеточное пространства. Введенный интравенозно инсулин достигает ликвора, не проходя через сплетения. Изолированные пиальная и эпендимальная поверхности вырабатывают жидкость, по химическому составу близкую к спинномозговой жидкости. Новейшие данные свидетельствуют о том, что арахноидальная мембрана участвует в экстрахориоидальном образовании ликвора. Существуют морфологические, а, вероятно, и функциональные различия между сосудистыми сплетениями боковых и IV желудочков. Считают, что около 70-85% спинномозговой жидкости появляется в сосудистых сплетениях, а остальное количество, то есть около 15-30%, — в мозговой паренхиме (мозговые капилляры, а также вода, образовавшаяся в процессе метаболизма).
Механизм образования ликвора (спинномозговой жидкости)
Согласно секреционной теории, ликвор является продуктом секреции сосудистых сплетений. Однако этой теорией нельзя объяснить отсутствие специфичного гормона и неэффективность воздействия некоторых стимуляторов и ингибиторов желез внутренней секреции на сплетения. По фильтрационной теории ликвор является обычным диализатом, или ультрафильтратом кровяной плазмы. Она объясняет некоторые общие свойства спинномозговой и интерстициальной жидкости.
Первоначально считалось, что это простая фильтрация. Позднее обнаружено, что для образования ликвора существенное значение имеет целый ряд биофизических и биохимических закономерностей:
- осмос,
- равновесие Донна,
- ультрафильтрация и др.
Биохимический состав ликвора наиболее убедительно подтверждает теорию фильтрации в целом, то есть то, что спинномозговая жидкость является только фильтратом плазмы. Ликвор содержит большое количество натрия, хлора и магния и низкое — калия, бикарбоната кальция фосфата и глюкозы. Концентрация этих веществ зависит от места получения спинномозговой жидкости, так как существует непрерывная диффузия между мозгом, экстрацеллюлярной жидкостью и ликвором при прохождении последнего через желудочки и подпаутинное пространство. Содержание воды в плазме около 93%, а в спинномозговой жидкости — 99%. Концентрационное соотношение ликвор/плазма в отношении большей части элементов существенно отличается от состава ультрафильтрата плазмы. Содержание белков, как было установлено при реакции Панди в ликворе, составляет 0,5% белков плазмы и изменяется с возрастом согласно формуле:
23,8 X 0,39 X возраст ± 0,15 г/л
Люмбальный ликвор, как показывает реакция Панди, содержит почти в 1,6 раза больше общих белков, чем желудочков, тогда как спинномозговая жидкость цистерн имеет в 1,2 раза больше общих белков, чем желудочков, соответственно:
- 0,06-0,15 г/л в желудочках,
- 0,15-0,25 г/л в мозжечково-продолговатомозговых цистернах,
- 0,20-0,50 г/л в люмбальном.
Считается, что высокий уровень белков в каудальной части образуется вследствие притока белков плазмы, а не в результате дегидратации. Эти различия не распространяются на все виды белков.
Соотношение ликвор/плазма для натрия — около 1,0. Концентрация калия, а по данным некоторых авторов, и хлора, уменьшается в направлении от желудочков к подпаутинному пространству, а концентрация кальция, напротив, увеличивается, тогда как концентрация натрия остается постоянной, хотя существуют и противоположные мнения. pH ликвора несколько ниже, чем pH плазмы. Осмотическое давление спинномозговой жидкости, плазмы и ультрафильтрата плазмы в обычном состоянии очень близки, даже изотоничны, что свидетельствует о свободном уравновешивании воды между этими двумя биологическими жидкостями. Концентрация глюкозы и аминокислот (например, глицина) очень низкая. Состав ликвора при изменениях концентрации плазмы остается почти постоянным. Так, содержание калия в спинномозговой жидкости остается в пределах 2-4 ммоль/л, тогда как в плазме его концентрация изменяется от 1 до 12 ммоль/л. С помощью гомеостазного механизма на постоянном уровне поддерживаются концентрации калия, магния, кальция, АК, катехоламинов, органических кислот и оснований, а также pH. Это имеет большое значение, так как изменения состава ликвора влекут за собой нарушения деятельности нейронов и синапсов ЦНС и изменяют нормальные функции мозга.
В результате развития новых методов исследования ликворной системы (вентрикуло-цистернальная перфузия in vivo, изолирование и перфузия сосудистых сплетений in vivo, экстракорпоральная перфузия изолированного сплетения, непосредственный забор жидкости из сплетений и ее анализ, контрастная радиография, определение направления транспорта растворителя и растворенных веществ через эпителий) возникла потребность рассмотрения вопросов, связанных с образованием ликвора.
Как следует рассматривать жидкость, образованную сосудистыми сплетениями? Как простой фильтрат плазмы, полученный в результате трансэпендимальных различий гидростатического и осмотического давления, или как специфичный сложный секрет ворсинчатых клеток эпендимы и других клеточных структур, возникший в результате затраты энергии?
Механизм ликворной секреции — довольно сложный процесс и хотя известны многие его фазы, все же еще остались нераскрытые звенья. Активный везикулярный транспорт, облегченная и пассивная диффузия, ультрафильтрация и другие виды транспорта играют определенную роль в образовании ликвора. Первым этапом в образовании спинномозговой жидкости является прохождение ультрафильтрата плазмы через капиллярный эндотелий, в котором отсутствуют уплотненные контакты. Под влиянием гидростатического давления в капиллярах, расположенных у основания хориоидальных ворсинок, ультрафильтрат поступает в окружающую соединительную ткань под эпителий ворсинок. Здесь определенную роль играют пассивные процессы. Следующий этап в образовании ликвора — это трансформирование поступающего ультрафильтрата в секрет, называемый ликвором. При этом большое значение имеют активные метаболические процессы. Иногда эти две фазы трудно отделить одну от другой. Пассивное всасывание ионов происходит с участием экстрацеллюлярного шунтирования в сплетения, то есть через контакты и латеральные межклеточные пространства. Кроме того, наблюдается пассивное проникновение через мембраны неэлектролитов. Происхождение последних во многом зависит от их растворимости в липидах/воде. Анализ данных свидетельствует о том, что проницаемость сплетений изменяется в очень широких пределах (от 1 до 1000*10-7 см/с; для сахаров — 1,6*10-7 см/с, для мочевины — 120*10-7 см/с, для воды 680*10-7 см/с, для кофеина — 432*10-7 см/с и т. д.). Вода и мочевина проникают быстро. Скорость их проникновения зависит от коэффициента липиды/вода, который может влиять на время проникновения через липидные мембраны этих молекул. Сахара проходят этот путь с помощью так называемой облегченной диффузии, которая показывает определенную зависимость от гидроксильной группы в молекуле гексозы. До настоящего времени отсутствуют данные об активном транспорте глюкозы через сплетения. Низкая концентрация сахаров в спинномозговой жидкости объясняется высокой скоростью метаболизма глюкозы в мозгу. Для образования ликвора большое значение имеют активные транспортные процессы против осмотического градиента.
Открытие Davson того факта, что движение Na+ от плазмы к ликвору однонаправленное и изотоничное с образованной жидкостью, стало оправдано при рассмотрении процессов секреции. Доказано, что натрий транспортируется активно и является основой процесса секреции спинномозговой жидкости из сосудистых сплетений. Опыты со специфичными ионными микроэлектродами показывают, что натрий проникает в эпителий благодаря существующему электрохимическому потенциальному градиенту, равному приблизительно 120 ммоль, через базо-латеральную мембрану эпителиальной клетки. После этого он поступает из клетки к желудочку против градиента концентрации через апикальную клеточную поверхность с помощью натриевого насоса. Последний локализован на апикальной поверхности клеток вместе с аденилциклоазотом и щелочной фосфатазой. Выделение натрия в желудочки происходит в результате проникновения туда воды вследствие осмотического градиента. Калий движется в направлении от ликвора к эпителиальным клеткам против градиента концентрации с затратой энергии и при участии калиевого насоса, расположенного также на апикальной стороне. Небольшая часть К+ после этого движется в кровь пассивно, вследствие электрохимического потенциального градиента. Калиевый насос связан с натриевым насосом, так как оба насоса имеют одинаковое отношение к уабаину, нуклеотидам, бикарбонатам. Калий перемещается только в присутствии натрия. Считают, что число насосов всех клеток составляет 3×106 и каждый насос осуществляет 200 перекачек в минуту.

1 — строма, 2 — вода, 3 — ликвор
В последние годы выявлена роль анионов в процессах секреции. Транспорт хлора, вероятно, осуществляется с участием активного насоса, но пассивное перемещение также наблюдается. Образование НСО3— из CO2 и Н2O имеет большое значение в физиологии ликвора. Почти все количество бикарбоната в спинномозговой жидкости образуется из CO2, а не переходит из плазмы. Этот процесс тесно связан с транспортом Na+. Концентрация HCO3— в процессе образования ликвора намного выше, чем в плазме, тогда как содержание Cl— низкое. Фермент карбоангидраза, который служит катализатором реакции образования и диссоциации угольной кислоты:

Этот фермент играет важную роль в секреции ликвора. Образующиеся протоны (Н+) обмениваются на поступающий в клетки натрий и переходят в плазму, а буферные анионы следуют за натрием в спинномозговой жидкости. Ацетазоламид (диамокс) является ингибитором этого фермента. Он существенно уменьшает образование ликвора или его ток, или то и другое. С введением ацетазоламида обмен натрия уменьшается на 50-100%, а скорость его непосредственно коррелирует со скоростью образования спинномозговой жидкости. Исследование новообразованного ликвора, взятого непосредственно из сосудистых сплетений, показывает, что он слегка гипертоничен вследствие активной секреции натрия. Это обусловливает осмотический водный переход от плазмы к ликвору. Содержание натрия, кальция и магния в спинномозговой жидкости несколько выше, чем в ультрафильтрате плазмы, а концентрация калия и хлора ниже. Вследствие сравнительно большого просвета хориоидальных сосудов можно допустить участие гидростатических сил в секреции ликвора. Около 30% этой секреции может быть не заторможено, это указывает на то, что процесс происходит пассивно, через эпендиму и зависит от гидростатического давления в капиллярах.
Уточнено действие некоторых специфичных ингибиторов. Уабаин ингибирует Na/K в зависимости от АТФ-азы и тормозит транспорт Na+. Ацетазоламид ингибирует карбоангидразу, а вазопрессин вызывает спазм капилляров. Морфологические данные детализируют клеточную локализацию части этих процессов. Иногда перенос воды, электролитов и других соединений в межклеточных хориоидных пространствах находится в состоянии коллапса (смотрите рисунок ниже). При ингибировании транспорта межклеточные пространства расширяются вследствие сжатия клеток. Рецепторы уабаина расположены между микроворсинками на апикальной стороне эпителия и обращены к ликворному пространству.

Segal и Роllау допускают, что образование ликвора можно разделить на две фазы (смотрите рисунок ниже). В первой фазе вода и ионы переносятся к ворсинчатому эпителию вследствие существования внутри клеток локальных осмотических сил, согласно гипотезе Diamond и Bossert. После этого во второй фазе ионы и вода переносятся, выходя из межклеточных пространств, в двух направлениях:
- в желудочки через апикальные уплотненные контакты и
- внутриклеточно и затем через плазматическую мембрану в желудочки. Эти трансмембранные процессы, вероятно, зависят от натриевого насоса.

1 — нормальное ликворное давление,
2 — повышенное ликворное давление
Ликвор в желудочках, мозжечково-продолговатомозговой цистерне и подпаутинном пространстве неодинаков по составу. Это свидетельствует о существовании экстрахориоидальных процессов обмена в ликворных пространствах, эпендиме и пиальной поверхности мозга. Это доказано для К+. От сосудистых сплетений мозжечково-продолговатомозговой цистерны концентрации К+, Са2+ и Mg2+ уменьшаются, в то время как концентрация Cl— увеличивается. Ликвор из подпаутинного пространства имеет более низкую концентрацию К+, чем субокципитальный. Сосудистая оболочка относительно проницаема для К+. Комбинацией активного транспорта в спинномозговой жидкости при полном насыщении и постоянной по объему секреции ликвора из сосудистых сплетений можно объяснить концентрацию этих ионов в только что образованной спинномозговой жидкости.
Резорбция и отток ликвора (спинномозговой жидкости)
- арахноидальных ворсинок (церебральных и спинальных);
- лимфатической системы;
- мозга (адвентиция мозговых сосудов);
- сосудистых сплетений;
- капиллярного эндотелия;
- арахноидальной мембраны.
Арахноидальные ворсинки считают местом дренажа ликвора, поступающего из субарахноидального пространства в синусы. Еще в 1705 г. Pachion описал арахноидальные грануляции, названные позднее его именем — пахионовы грануляции. Позже Key и Retzius указывали на значение арахноидальных ворсинок и грануляций для оттока ликвора в кровь. Кроме того, несомненно, что в резорбции спинномозговой жидкости участвуют мембраны, соприкасающиеся с ликвором, эпителий оболочек цереброспинальной системы, мозговая паренхима, периневральные пространства, лимфатические сосуды и периваскулярные пространства. Участие этих дополнительных путей невелико, но они приобретают большое значение, когда главные пути затронуты патологическими процессами. Самое большое количество арахноидальных ворсинок и грануляций находится в зоне верхней сагиттальной пазухи. В последние годы получены новые данные относительно функциональной морфологии арахноидальных ворсинок. Их поверхность образует один из барьеров для оттока ликвора. Поверхность ворсинок изменчива. На их поверхности находятся веретенообразные клетки 40-12 мкм длиной и 4-12 мкм толщиной, в центре находятся апикальные выпуклости. Поверхность клеток содержит многочисленные маленькие выпуклости, или микроворсинки, и соседние с ними пограничные поверхности имеют неправильные очертания.
Ультраструктурные исследования показывают, что поверхности клеток поддерживают поперечные базальные мембраны и субмезотелиальная соединительная ткань. Последняя состоит из коллагеновых волокон, эластичной ткани, микроворсинок, базальной мембраны и мезотелиальных клеток с длинными и тонкими цитоплазматическими отростками. Во многих местах отсутствует соединительная ткань, вследствие чего образуются пустые пространства, которые находятся в связи с межклеточными пространствами ворсинок. Внутренняя часть ворсинок образована соединительной тканью, богатой клетками, ограждающими лабиринт от межклеточных пространств, которые служат продолжением арахноидальных пространств, содержащих ликвор. Клетки внутренней части ворсинок имеют различные формы и ориентацию и похожи на клетки мезотелия. Выпуклости близкостоящих клеток связаны между собой и образуют единое целое. Клетки внутренней части ворсинок имеют хорошо выраженный сетчатый аппарат Гольджи, цитоплазматические фибриллы и пиноцитозные везикулы. Между ними иногда находятся «блуждающие макрофаги» и различные клетки лейкоцитарного ряда. Так как эти арахноидальные ворсинки не содержат кровеносных сосудов и нервов, считают, что они питаются спинномозговой жидкостью. Поверхностные мезотелиальные клетки арахноидальных ворсинок образуют с близлежащими клетками непрерывную мембрану. Важным свойством этих мезотелиальных клеток, покрывающих ворсинки, является то, что они содержат одну или несколько гигантских вакуолей, вздутых в направлении апикальной части клеток. Вакуоли соединены с мембранами и обычно пусты. Большая часть вакуолей вогнута и непосредственно связана с ликвором, находящимся в субмезотелиальном пространстве. У значительной части вакуолей базальные отверстия больше апикальных и эти конфигурации интерпретируют как межклеточные каналы. Изогнутые вакуольные трансцеллюлярные каналы выполняют функцию одностороннего клапана для оттока ликвора, то есть в направлении базиса к верхушке. Структура этих вакуолей и каналов хорошо изучена с помощью меченых и флуоресцентных веществ, вводимых чаще всего в мозжечково-продолговатомозговую цистерну. Трансцеллюлярные каналы вакуолей представляют собой динамическую систему пор, которая играет основную роль в резорбции (оттока) ликвора. Считают, что некоторая часть из предполагаемых вакуольных трансцеллюлярных каналов, в сущности, является расширенными межклеточными пространствами, которые также имеют большое значение для оттока ликвора в кровь.
Еще в 1935 году Weed на основании точных опытов установил, что часть ликвора оттекает через лимфатическую систему. В последние годы появился ряд сообщений о дренаже спинномозговой жидкости через лимфатическую систему. Однако эти сообщения оставили открытым вопрос о том, какое количество ликвора абсорбируется и какие механизмы в этом участвуют. Через 8—10 ч после введения в мозжечково-продолговатомозговую цистерну окрашенного альбумина или меченых белков от 10 до 20% этих веществ можно обнаружить в лимфе, образующейся в шейном отделе позвоночника. При увеличении внутрижелудочкового давления дренаж через лимфатическую систему усиливается. Ранее предполагалось, что существует резорбция ликвора через капилляры мозга. При помощи компьютерной томографии установлено, что перивентрикулярные зоны пониженной плотности часто обусловлены поступлением ликвора экстрацеллюлярно в ткани мозга, особенно при увеличении давления в желудочках. Спорным остается вопрос о том, является ли поступление большей части спинномозговой жидкости в мозг резорбцией или последствием дилатации. Наблюдается вытекание ликвора в межклеточное мозговое пространство. Макромолекулы, которые вводятся в вентрикулярную спинномозговую жидкость или субарахноидальное пространство, быстро достигают внеклеточного мозгового пространства. Сосудистые сплетения считают местом оттока ликвора, так как они окрашиваются после введения краски при увеличении ликворного осмотического давления. Установлено, что сосудистые сплетения могут резорбировать около 1/10 секретированного ими ликвора. Этот отток чрезвычайно важен при высоком внутрижелудочковом давлении. Спорными остаются вопросы абсорбции ликвора через капиллярный эндотелий и арахноидальную мембрану.
Механизм резорбции и оттока ликвора (спинномозговой жидкости)
Для резорбции ликвора имеет значение целый ряд процессов: фильтрация, осмос, пассивная и облегченная диффузия, активный транспорт, везикулярный транспорт и другие процессы. Отток ликвора можно характеризовать как:
- однонаправленное просачивание через арахноидальные ворсинки посредством клапанного механизма;
- резорбция, которая не является линейной и требует определенного давления (обычно 20-50 мм вод. ст.);
- своеобразный пассаж из спинномозговой жидкости в кровь, но не наоборот;
- резорбция ликвора, уменьшающаяся, когда общее содержание белка увеличивается;
- резорбция с одинаковой скоростью для молекул различных размеров (например, молекул маннитола, сахарозы, инсулина, декстрана).
Скорость резорбции спинномозговой жидкости зависит в значительной степени от гидростатических сил и является относительно линейной при давлении в широких физиологических пределах. Существующая разница в давлении между ликвором и венозной системой (от 0,196 до 0,883 кПа) создает условия для фильтрации. Большое различие в содержании белка в этих системах определяет значение осмотического давления. Welch и Friedman предполагают, что арахноидальные ворсинки функционируют как клапаны и определяют движение жидкости в направлении от ликвора к крови (в венозные синусы). Размеры частиц, которые проходят через ворсинки, различны (коллоидное золото размером 0,2 мкм, полиэфирные частички — до 1,8 мкм, эритроциты — до 7,5 мкм). Частички с большими размерами не проходят. Механизм оттока ликвора через различные структуры различен. В зависимости от морфологической структуры арахноидальных ворсинок существует несколько гипотез. Согласно закрытой системе, арахноидальные ворсинки покрыты эндотелиальной мембраной и между клетками эндотелия находятся уплотненные контакты. Вследствие наличия этой мембраны резорбция ликвора совершается с участием осмоса, диффузии и фильтрации низкомолекулярных веществ, а для макромолекул — путем активного транспорта через барьеры. Однако прохождение некоторых солей и воды остается свободным. В противоположность этой системе существует открытая система, согласно которой в арахноидальных ворсинках имеются открытые каналы, связывающие паутинную оболочку с венозной системой. Эта система предполагает пассивное прохождение микромолекул, в результате чего абсорбция спинномозговой жидкости полностью зависит от давления. Tripathi предложил еще один механизм абсорбции ликвора, который, в сущности, является дальнейшим развитием первых двух механизмов. Помимо последних моделей, существуют еще динамические трансэндотелиальные вакуолизационные процессы. В эндотелии арахноидальных ворсинок временно образуются трансэндотелиальные или трансмезотелиальные каналы, через которые ликвор и его составные частицы вытекают из субарахноидального пространства в кровь. Эффект давления при этом механизме не выяснен. Новые исследования подкрепляют эту гипотезу. Считают, что с увеличением давления число и размеры вакуолей в эпителии возрастают. Вакуоли с размерами больше 2 мкм встречаются редко. Комплексность и интеграция уменьшаются при больших различиях в давлении. Физиологи считают, что резорбция ликвора является пассивным, зависящим от давления процессом, который происходит через поры, размеры которых больше размеров молекул протеинов. Спинномозговая жидкость проходит от дистального субарахноидального пространства между клетками, образующими строму арахноидальных ворсинок и достигает субэндотелиального пространства. Однако эндотелиальные клетки пиноцитозно активны. Прохождение ликвора через эндотелиальный слой является также активным трансцеллюлозным процессом пиноцитоза. Согласно функциональной морфологии арахноидальных ворсинок, прохождение спинномозговой жидкости осуществляется через вакуольные трансцеллюлозные каналы в одном направлении от базиса к верхушке. Если давление в подпаутинном пространстве и синусах одинаковое, арахноидальные разрастания находятся в состоянии коллапса, элементы стромы плотные и эндотелиальные клетки имеют суженные межклеточные пространства, местами пересеченные специфическими клеточными соединениями. Когда в субарахноидальном пространстве давление повышается только до 0, 094 кПа, или 6-8 мм вод. ст., разрастания увеличиваются, клетки стромы отделяются одна от другой и эндотелиальные клетки выглядят меньшими по объему. Межклеточное пространство расширено и клетки эндотелия проявляют повышенную активность к пиноцитозу (смотрите рисунок ниже). При большой разнице в давлении изменения более выражены. Трансцеллюлярные каналы и расширенные межклеточные пространства позволяют прохождение ликвора. Когда арахноидальные ворсинки находятся в состоянии коллапса, проникновение составных частиц плазмы в спинномозговую жидкость невозможно. Для резорбции ликвора имеет значение также микропиноцитоз. Прохождение молекул протеина и других макромолекул из спинномозговой жидкости субарахноидального пространства зависит в известной степени от фагоцитарной активности арахноидальных клеток и «блуждающих» (свободных) макрофагов. Вряд ли, однако, чтобы клиренс этих макрочастичек осуществлялся только путем фагоцитоза, так как это достаточно продолжительный процесс.

1 — арахноидальные ворсинки, 2 — хориоидальное сплетение, 3 — субарахноидальное пространство, 4 — оболочки мозга, 5 — боковой желудочек.
В последнее время все больше становится сторонников теории активной резорбции ликвора через сосудистые сплетения. Точный механизм этого процесса не выяснен. Однако предполагают, что вытекание спинномозговой жидкости происходит в сторону сплетений из субэпендимального поля. После этого через фенестрированные ворсинчатые капилляры ликвор поступает в кровь. Эпендимальные клетки с места резорбционных транспортных процессов, то есть специфичные клетки, являются посредниками для переноса веществ из вентрикулярного ликвора через ворсинчатый эпителий в кровь капилляров. Резорбция отдельных составных частей спинномозговой жидкости зависит от коллоидного состояния вещества, его растворимости в липидах/воде, отношения к специфичным транспортным белками и т. д. Для переноса отдельных компонентов существуют специфичные транспортные системы.
Скорость образование ликвора и резорбции спинномозговой жидкости
Методы исследования скорости образование ликвора и резорбции спинномозговой жидкости, которые использовались до настоящего времени (продолжительный люмбальный дренаж; вентрикулярный дренаж, используемый также для лечения гидроцефалии; измерение времени, необходимого для восстановления в ликворной системе давления, после истечения спинномозговой жидкости из субарахноидального пространства), подвергались критике за то, что они были нефизиологичными. Метод вентрикулоцистернальной перфузии, введенный Pappenheimer и соавторами, был не только физиологичным, но и позволял одновременно производить оценку образования и резорбции ликвора. Скорость образования и резорбции спинномозговой жидкости определялась при нормальном и патологическом давлении спинномозговой жидкости. Образование ликвора не зависит от непродолжительных изменений вентрикулярного давления, отток его линейно связан с ним. Секреция ликвора уменьшается при продолжительном повышении давления в результате изменений в хориоидальном кровотоке. При давлении ниже 0,667 кПа резорбция равна нулю. При давлении между 0,667 и 2,45 кПа, или 68 и 250 мм вод. ст. соответственно, скорость резорбции спинномозговой жидкости прямо пропорциональна давлению. Cutler и соавторы изучали эти явления у 12 детей и установили, что при давлении 1,09 кПа, или 112 мм вод. ст., скорость образования и скорость оттока ликвора равны (0,35 мл/мин). Segal и Pollay утверждают, что у человека скорость образования спинномозговой жидкости достигает 520 мл/мин. Еще мало известно об эффекте воздействия температуры на образование ликвора. Экспериментально остро вызванное повышение осмотического давления тормозит, а понижение осмотического давления усиливает секрецию ликвора. Неврогенное стимулирование адренергических и холинергических волокон, которые иннервируют хориоидальные кровеносные сосуды и эпителий, имеют различное действие. При стимулировании адренергических волокон, которые исходят из верхнего шейного симпатического узла, ток ликвора резко уменьшается (почти на 30%), а денервирование усиливает его на 30%, не изменяя хориоидальный кровоток.
Стимулирование холинергического пути увеличивает образование ликвора до 100%, не нарушая хориоидальный кровоток. В последнее время выясняется роль цикличного аденозинмонофосфата (цАМФ) в прохождении воды и растворенных веществ через клеточные мембраны, в том числе и влияние на сосудистые сплетения. Концентрация цАМФ зависит от активности аденилциклазы, фермента, который катализирует образование цАМФ из аденозинтрифосфата (АТФ) и активности его метаболизирования до неактивного 5-АМФ с участием фосфодиэстеразы, или присоединения к нему ингибиторной субъединицы специфичной протеинкиназы. цАМФ действует на ряд гормонов. Холерный токсин, который является специфичным стимулятором аденилциклазы, катализирует образование цАМФ, при этом наблюдается пятикратное увеличение этого вещества в сосудистых сплетениях. Ускорение, вызванное холерным токсином, можно блокировать препаратами из группы индометацина, которые являются антагонистами по отношению к простогландинам. Спорным является вопрос, какие специфичные гормоны и эндогенные агенты стимулируют образование спинномозговой жидкости по пути к цАМФ и каков механизм их действия. Имеется обширный список лекарств, которые влияют на образование спинномозговой жидкости. Некоторые лекарственные препараты воздействуют на образование ликвора как препятствующие метаболизму клеток. Динитрофенол влияет на окислительное фосфорилирование в сосудистых сплетениях, фуросемид — на транспорт хлора. Диамокс уменьшает скорость образования спинномозговой путем торможения карбоангидразы. Он также вызывает преходящее повышение внутричерепного давления, освобождая CO2 из тканей, следствием чего является увеличение мозгового кровотока и объема крови мозга. Сердечные гликозиды тормозят Na- и К-зависимость АТФ-азы и уменьшают секрецию ликвора. Глико- и минералокортикоиды почти не влияют на обмен натрия. Увеличение гидростатического давления действует на процессы фильтрации через капиллярный эндотелий сплетений. При повышении осмотического давления путем введения гипертонического раствора сахарозы или глюкозы образование ликвора уменьшается, а при снижении осмотического давления введением водных растворов — увеличивается, так как эта связь почти линейная. При изменении осмотического давления введением 1% воды скорость образования спинномозговой жидкости нарушается. При введении гипертонических растворов в терапевтических дозах осмотическое давление увеличивается на 5-10%. Внутричерепное давление значительно больше зависит от церебральной гемодинамики, чем от скорости образования спинномозговой жидкости.
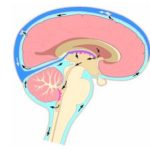
Циркуляция ликвора (спинномозговой жидкости)

1 — спинальные корешки, 2 — хориоидальные сплетения, 3 — хориоидальные сплетения, 4 — III желудочек, 5 — хориоидальное сплетение, 6 — верхняя сагиттальная пазуха, 7 — арахноидальная гранула, 8 — боковой желудочек, 9 — полушарие головного мозга, 10 — мозжечок.
Циркуляция ликвора (спинномозговой жидкости) изображена на рисунке выше.